
一、己糖磷酸旁路
在红细胞中并不是全部葡萄糖的代谢都要经过EMP途径,约5%~10%的葡萄糖经氧化途径。氧化结果,葡萄糖第一碳氧化为二氧化碳,并生成戊糖磷酸,同时它与耦联的NADP+被还原为NADPH。戊糖经一系列分子重排变成GA3P和F6P。它们都是EMP的中间产物,因此又重新加入EMP代谢。
用放射性核素14C标记,葡萄糖第一碳变成CO2,其他5碳重循环量很小,但在有亚甲蓝或维生素C的参加下可增加到50%,说明这个旁路代谢有很大的伸缩性。主要作用是保证红细胞不受氧化作用所损害。旁路直接或间接与NADPH和GSH系统耦联其作用就是为了把H2O2还原为H2O。因此葡萄糖虽然只有5%~10%经过这个旁路,却是红细胞存活所必不可少。不论先天或药物性的旁路堵塞,常常与各种血液病相联系。蚕豆病是其中突出的例子。
葡萄糖经过HMP不需ATP的参加而把戊糖磷酸化为R5P。R5P在红细胞内又可合成为磷酸戊糖焦磷酸(PRPP),它可参与EMP和红细胞代谢的修复,尤其在延长血液保存期的工作中很受重视。己糖磷酸旁路见下图。
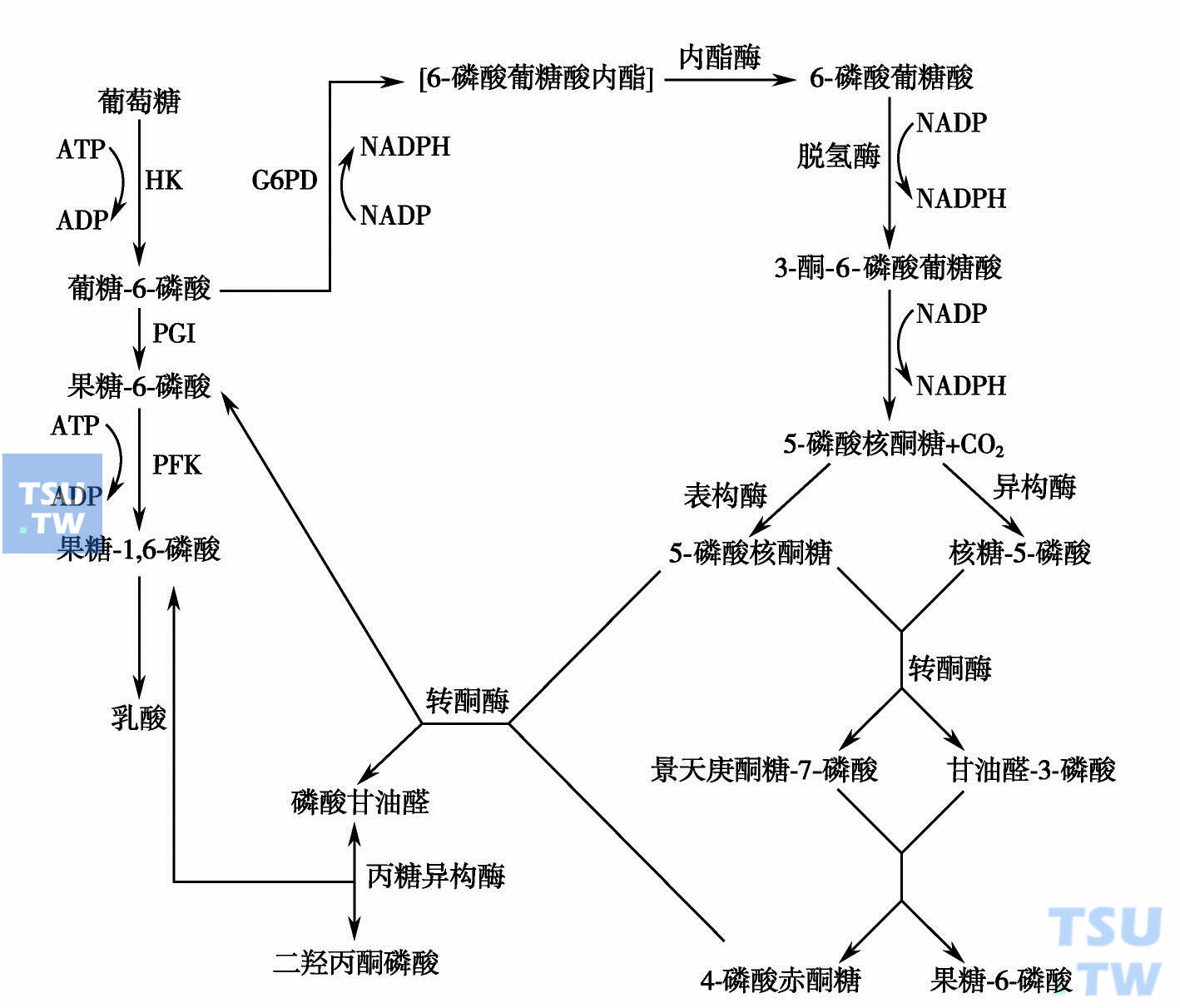
己糖磷酸旁路(引自:邓家栋.临床血液学,2001:386)
二、己糖磷酸旁路途径与红细胞的氧化还原反应红细胞经常接触高氧压,容易被氧化。HMP是抗氧化基地和修复途径,保持谷胱甘肽和NADPH的还原型。通过多级氧化还原系统的耦联,使一些被氧化的化合物重新还原,以恢复其生理作用。各级耦联作用都有相应还原酶的参加,具体情况参见下图:
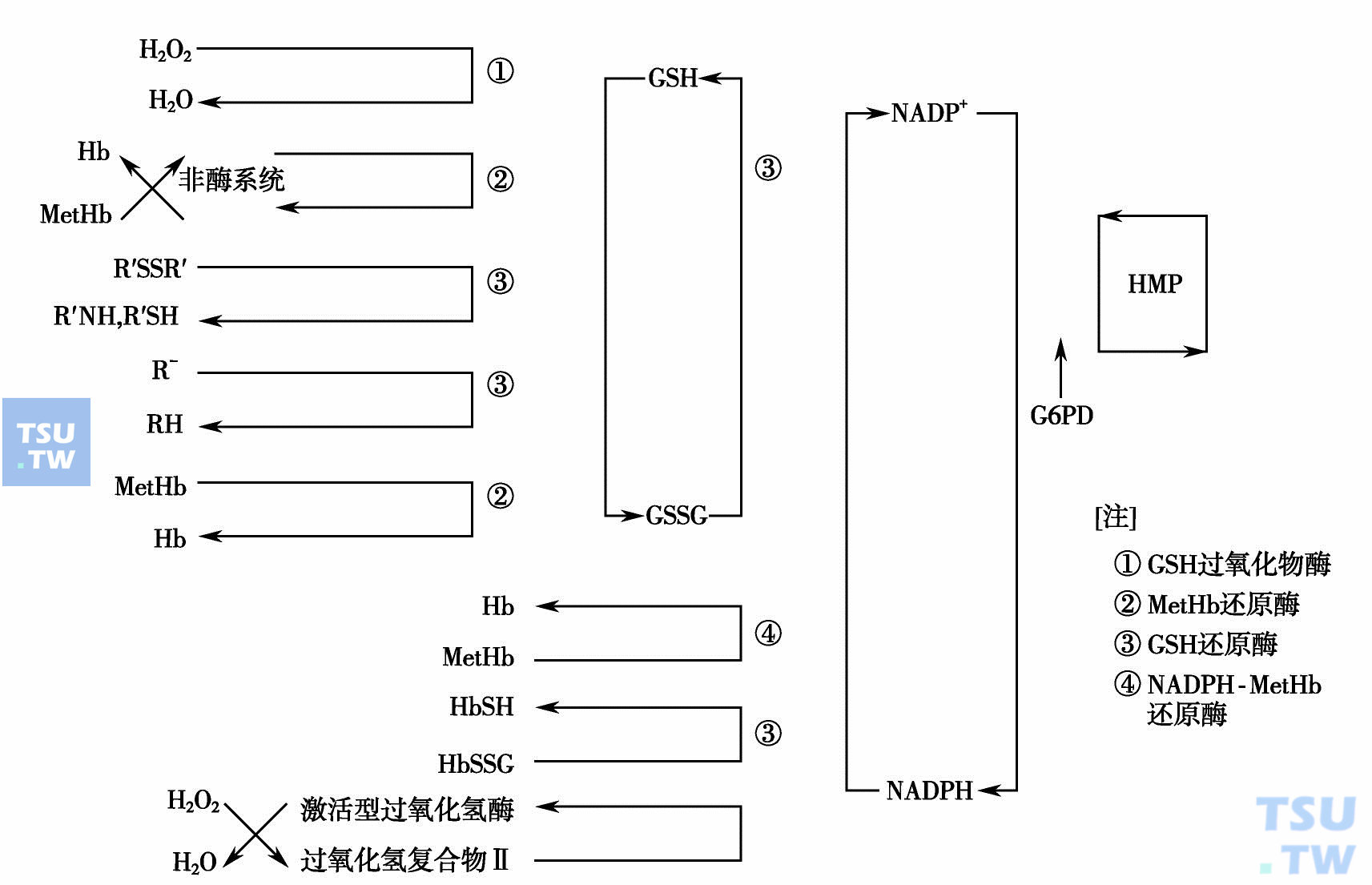
己糖磷酸旁路和红细胞的氧化还原反应(多级氧化还原反应)(引自:邓家栋.临床血液学,2001:387)
谷胱甘肽的存活时间只有4天,它可自由进出红细胞膜;红细胞可合成这个化合物。尽管有过氧化氢酶和GSH过氧化物酶对在正常情况下代谢产生或药物引起的低浓度H2O2的解毒,GSH还很重要。Teitel(1968)曾经发现把GSH封闭后出现的后果是增加红细胞的破裂和加速ATP的消耗,膜-SH被氧化,Heinz小体的形成;最后延长红细胞通过模拟脾筛过滤器的时间。GSSG虽可还原为GSH,但一旦生成就会透过细胞膜,有保护红细胞的作用,因为GSSG存在对HK能起抑制作用。
谷胱甘肽还原酶含有核黄素,因此它的活性受食物中核黄素含量的影响。此酶在4℃和-30℃都很稳定,红细胞缺乏此酶,HMP活力几乎不受影响。
NADPH-MetHb还原酶(心肌黄酶Ⅱ)比NADHMetHb还原酶(心肌黄酶Ⅰ)在最适条件下活力小10倍,说明MetHb的生成不大牵涉HMP。NADPH 对MetHb的还原被非酶氧化还原系统所加强。NADPH可激活过氧化氢酶,后者的去激活将造成HMP代谢速度的加快。因此,旁路的活力首先决定于NADPH和GSH的可用性;同时也取决于氧化威胁的程度。一个红细胞缺过氧化氢的人,在氧化威胁下,HMP代谢可高达正常的20倍。这是由于缺乏过氧化氢酶迫使GSH过氧化物酶作用的结果。旁路活动比单纯克服氧威胁需要还大,其可能性是:①通过R5P增加腺苷酸的合成;②增加EMP后半的中间产物。两个途径对G6P竞争;对HMP最适pH要低些,即酸中毒有利于HMP的活动。pH7. 1 和7. 4相比对HMP活力无大区别,而pH7. 1时EMP活力只有50%。当NADPH-MetHb还原酶不足以应付时,则需依赖NADH,NAD/NADH的比例则升高,代谢加快,2,3-DPG含量也增加。另外过氧化氢酶活力下降,MetHb堆积将导致NADH-MetHb还原酶活性增加,NAD/NADH也要增加,2,3-DPG也可能增加。
旁路酶G-6-PD:是旁路代谢控制酶,存在单体和四聚体。它的活力受GSSG、ADP、Mg2+和本身浓度的影响。G-6-PD缺陷的基因在性染色体上,为伴性不完全显性遗传。
转酮酶(TK)和转醛酶(TA):两者活力的降低将等分子的影响HMP旁路的活力。赤鲜糖-4-磷酸(E4P)和核酮糖-5-磷酸(Xa-5-P)变成F6P和GA-3-P这个反应步骤很重要。少量E4P将抑制G6P变成F6P,同时抑制TK。E4P含量增加将停止HMP的代谢,因此降低C-2再循环进而降低整个红细胞代谢。
总之,虽然HMP对葡萄糖代谢在量上是次要的,但它的充分传输对红细胞的存活与EMP同样重要。旁路有3个主要功能,即维持红细胞的还原能力、核苷酸的再生(尤其PRPP)和提供EMP后半段代谢的底物。但在正常状况下,旁路支付给EMP和对付多种氧化性威胁的分量不详。HMP在红细胞代谢中的功能上未完全了解。
三、氧化还原系统
红细胞内利用葡萄糖的5%~10%通过磷酸戊糖通路代谢,为红细胞提供另一种还原力(NADPH),NADPH在红细胞氧化还原系统中发挥重要作用,具有保护膜蛋白、血红蛋白及酶蛋白的巯基不被氧化,还原高铁血红蛋白等多种功能。
人的红细胞中谷胱甘肽含量甚多(2×10-3mol/ L)。正常时,谷胱甘肽几乎全部是还原型(GSH)。谷胱甘肽可在红细胞内合成,它的合成原料为谷氨酸、半胱氨酸和甘氨酸。其合成过程为:谷氨酸与半胱氨酸在ATP和γ-谷氨酸半胱氨酸合成酶的参与下,缩合成二肽γ-谷氨酸半胱氨酸,后者与甘氨酸在ATP和谷胱甘肽合成酶的参与下缩合成谷胱甘肽。
GSH具有重要的生理功能。它能通过GSH过氧化物酶还原体内生成的过氧化氢,以消除后者对血红蛋白、酶和膜蛋白上的巯基(—SH)的氧化作用,维持其生物活性。细胞内可自发生成少量超氧阴离子,同时感染时的白细胞吞噬作用亦可产生超氧阴离子,可被超氧化物歧化酶(SOD)催化生成过氧化氢。在此反应中,GSH还原H2O2成水,而自身氧化为氧化型谷胱甘肽(GSSG)。
GSSG可经红细胞中谷胱甘肽还原酶的催化,利用磷酸戊糖旁路所生成的NADPH,重新还原成GSH。
当磷酸戊糖旁路中最常见的6-磷酸葡萄糖脱氢酶(G-6-PD)缺失时,患者容易发生溶血现象。该病患者的红细胞中,NADPH生成受阻,因此GSH含量减少。由于GSSG和GSS-Hb增多致使Hb解链和变性,形成Heinz小体。Heinz小体形成后,红细胞表面电荷和形态可塑性均发生相应的改变,如红细胞膜变得僵硬易破碎,不易通过脾窦小孔,因阻留而被破坏导致溶血。
